一直是個謎?
脈絡很清晰,只是你不信,甚至不想去了解罷了。
生物學發展到21世紀的今天,無論古生物學(化石鏈)、遺傳學、組胚學、解剖學、比較生物學、系統發育學、分子生物學/人類學,還是神經科學,都對人類起源有著強有力的支持。
5.18億年前——最早的脊索動物(或半索動物),澄江生物群的雲南蟲繁榮。

5.15億年前——在奇蝦制霸下瑟瑟發抖的皮卡蟲,構造簡單接近文昌魚。

僅僅是幾厘米長的底棲頭索動物,以浮遊生物爲生[1]。爲了在水中呼吸,擁有幾十上百個的鰓裂。
在早期泛節肢動物的制霸下,瑟瑟發抖。

5.1億年前——在不斷被淘汰的過程中,活下來的祖先逐漸有了脊椎,以及一節一節的肌肉,化身成昆明魚/海口魚[2]。
無颌的嘴,只能吸食食物。

原始圓口類,對標現今的盲鳗、七鰓鳗。
從原始魚類開始,鰓弓不斷減少,進化成其他器官。
作爲最早的脊椎動物,它們已經鋪墊了後世所有脊椎動物的神經系統:
它們的大腦分成大腦、間腦、中腦、小腦、延腦五個部分。

雖然因爲圖片僞造,海克爾胚胎重演律在生物學史上頗具爭議,但所有脊索動物胚胎,大腦發育的確會經曆相似的過程,雖然算不上真正的“重演"。當初海克爾的僞造,正是刻意抹除了絕大部分的差異。
人類胚胎發育過程中,原始的腦泡會先發育成三個,然後再發育成5個:

4.8億年前——隨著進入奧陶紀後(4.8~4.4億年前),整個原始海洋都變成了一片修羅場。
早期海洋的超級霸主——板足鲎(海蠍子)大約在4.7億年前登場。
它們很快發展爲龐大的家族,足足縱橫了海洋上億年,一直到泥盆紀才開始衰退[3]。

海蠍子憑借鋒利的大型節肢,在原始海洋,可謂是所向披靡。
在如此殘酷海洋裏,弱小的早期脊椎動物簡直就是行走的食物。
我們的祖先被大量捕食滅絕,最終那些偶然進化出骨質甲胄的才得以苟延殘喘,並發展出了早期無颌魚中比較繁榮的大家族——甲胄魚。

雖然盔甲可以爲它們提供保護,但它們的頭也與軀甲完全連爲一體,再加上過于笨重的身軀讓它們行動緩慢,且沒有颌反擊,如果遇到能咬穿它們盔甲的掠食者,它們唯一能做的就是任之宰割。
4.4億年前——奧陶紀大滅絕,史前動物大洗牌。
但依舊還有著長到1米多長的秀山恐鲎,便對它們展開了無情的屠殺。
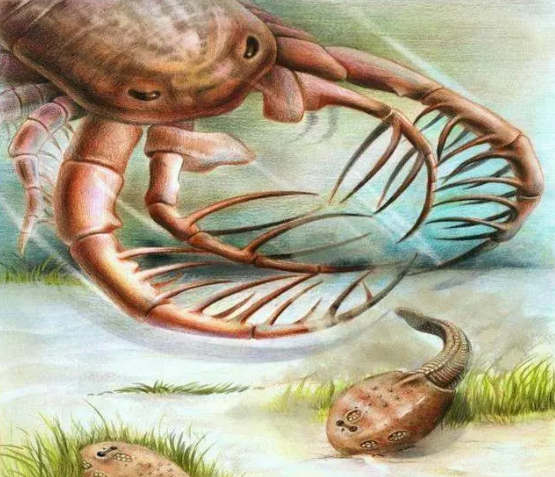
4.3億年前——初始全颌魚[4]。
終于,一個偉大的進化革命出現了。魚類第一次擁有了下颌和舌頭。

遺傳學證據表明,下颌起源的最早時間,可上溯到奧陶紀晚期,也即4.5億年前[5][6]。
去年中科院幾篇有颌魚起源的重磅1天4 篇 Nature中,發現了更原始的有颌魚——奇迹秀山魚[7]。
初始全颌魚犧牲了原本用于呼吸的第一鰓弓,演化出了颌。

颌出現過後,頭部可以靈活轉動,不僅它們的攝食能力變強了,有了反抗天敵的可能,而且還帶動了身體各個方面的進化。
它們通過不斷折疊鳍褶[8],進化出了完全的偶鳍,運動能力大大提升。
不僅能從天敵口中逃脫,還能更迅猛地捕食小型動物。
攝食和運動能力的加強,令早期魚類逐漸大型化,大腦也變得更加發達。
隨著神經纖維的增長,爲了保證神經信號的傳輸速度,促使了大腦高速信號通道——髓鞘的出現。

4.15億年前——甲胄魚。颌是如此重要,甚至當甲胄魚進化爲有颌、有完全偶鳍的盾皮魚後,完成了徹底的逆襲,徹底主宰了志留紀和泥盆紀。
海蠍子開始衰落,出現可達10米的鄧氏魚稱霸海洋。

4億年前——夢幻鬼魚,是最早的硬骨魚,此時鲨魚已經出現。

此時魚類的腦神經已經變得越來越發達,中腦爲主要的神經中樞。隨著感官的細化,也有了10對腦神經,爲後來神經系統的複雜演化奠定了基礎:

3.8億年前——肉鳍魚,魚鳍中出現中軸骨,魚鳍變得更加強壯有力。

第4對鰓弓和第6對鰓弓進化成了咽喉部位的軟骨、肌肉,以及相關神經。
大名鼎鼎的喉返神經,便是第6鰓弓進化而來的。

3.75億年前——提塔利克魚開始進軍陸地,魚鳍中已出現骨骼。


鰓逐漸全部退化,進化出了新的呼吸器官肺,感知水流的鼻孔演化成了呼吸的通道。
鰓弓進行上的殘留,也讓人類遭受一些莫名的病痛,例如下颌三叉神經痛,偏頭痛也而可能和相關神經有關。包括打嗝在內,也可能是進化爲兩棲動物時産生的。
至于耳盲管之類的進化BUG,出現發炎時,甚至可能危及生命。

雖然鰓對耳朵進化的貢獻很大,但它主要也在于優化聽覺(中耳三塊聽小骨——镫骨、砧骨和錘骨,具有傳遞和放大聲音的作用),而對聽覺的存在並沒有起到決定性的作用。
因爲魚類已經有了原始的耳朵和基本的聽覺。

這些決定了人類內耳的起源。
3.74億年前——希望螈,魚鳍上的鳍條退化,軟骨內成骨逐漸進化成指骨,于是四肢出現了,與人類手指有對應關系。



泥盆紀後期滅絕事件,海洋節肢動物衰退,陸地節肢動物興起。
3.67億年前——魚石螈,四肢關節靈活,開始陸地活動,手指減小到6~7根。

3.4億年——兩棲動物離片椎類出現,現今所有滑體兩棲類、爬行、鳥類、哺乳動物共同祖先。

兩棲動物的大腦比起魚類來說複雜不了多少。
但出現了植物性神經,以及原腦皮:

兩棲類的大腦,和人類的腦幹以及小腦部分極其的相似:
這是人類的腦幹和小腦:
植被的繁榮,帶來陸生物種的繁榮。
早期四足動物,先後演化出原腦皮層、古腦皮層、周圍舊皮層,以及中間皮層。
3.18億年前——動物的卵裏有了羊膜結構,分化出了蜥形綱與合弓綱。
現今的爬行類、鳥類,以及古生物學史上的恐龍,都源于蜥形綱;而合弓綱先發展出類哺乳動物,後發展出現代哺乳動物。
它們的端腦發達,成了大腦的主要結構,分爲三層。不僅增大的紋狀體,也令大腦體積越來越大,甚至第一次出現了新腦皮(新皮層),但此時的新皮層極其的原始。
在後來的演化中,新皮層逐漸演化成了處理各類信息的中樞。3.05億年前——雨林崩潰事件,森林分裂,哺乳動物祖先(類哺乳動物)迎來大發展。
3億年前——合弓綱繁榮,進化出盤龍類。

2.9億年前——似哺乳爬行動物繁榮,盤龍類進化出哺乳動物祖先獸孔類,水龍獸爲代表。

2.75億年前——基龍和異齒龍出現,與類哺乳動物相伴。
2.51億年前——史上最慘烈的滅絕事件,二疊紀大滅絕。
2.5億年前——類哺乳動物衰退,僅剩小型種類。
2.35億年前——最早的恐龍出現,恐龍時代開始。
恐龍和鳥類,大腦結構和變溫爬行動物大同小異,爲了適應飛翔,小腦變得更加發達,紋狀體多了上紋狀體。大腦主要結構依舊是三層的舊皮層,新皮層並未明顯的發展。
鳥類大腦與人類大腦的對比關系:

類哺乳動物(合弓綱)這一支,下丘腦、海馬體以及杏仁核則得到了充分發育,紋狀體則朝著基底神經核方向演化。
現代哺乳動物的大腦,則是在類哺乳動物大腦的基礎上發育而來的。
2.05億年前——著名的早期哺乳動物摩爾根獸出現。

2億年前——三疊紀末大滅絕。
1.95億年前——吳氏巨顱獸——最早的哺乳動物之一,出現比較發達的嗅覺和觸覺,出現了比較發達的新皮層。


1.7億年前——我們和袋鼠擁有共同的祖先。
1.48億年前——真獸下綱(包括胎盤類動物)和有袋動物分道揚镳。
1.35億年前——我們的祖先,有胎盤類哺乳動物攀援始祖獸出現了,但它們只有25克左右。

如果說,從魚類到哺乳動物,大腦的演化是不斷地在地基的基礎上加高樓層:

那麽,從低等哺乳動物到人類大腦的演化過程,就是在進行精裝修。
和鳥類進化紋狀體不同,哺乳動物走上了折疊新皮層的演化之路。相對來說,對于種內、種間競爭更大的物種,新皮層折疊的溝回也就越多越深。
被子植物的出現,以及繁榮,形成豐富多樣的現代叢林,也爲哺乳動物的發展提供豐富的生態環境。
8500萬年前——勞亞獸總目和靈長總目分道揚镳。

6500萬年前——白垩紀大滅絕,恐龍終,最早似靈長類更猴出現。

6000萬年前——普爾加托裏猴,酷似松鼠的早期靈長類。
它們此時的新皮層溝回應該還比較少,接近老鼠大腦的光滑度。

靈長總目這一支,從老鼠到猴子再到人類,大腦的整個演化過程,幾乎就是新皮層更加複雜化的折疊過程:

4500萬年前——蒙古安氏中獸爲代表的古食肉目稱霸大陸。最早的高級靈長動物中華曙猿出現,然而它們只有50克到150克之間。


3500萬年前——埃及猿出現,舊世界猴(如猕猴)和新世界猴的共同祖先,食肉目(貓狗祖先)開始崛起。

2500萬年前——舊世界猴進化出古猿。
1900萬年前——原康修爾猿,長臂猿與我們分道揚镳。
因爲抓握能力的強化,智商比起其他靈長類得到了很大的提升。

1200萬年前——西瓦古猿,紅毛猩猩與我們分道揚镳。
作爲人類和紅毛猩猩的共同祖先,已經能開始用簡單的現成工具(木棍什麽的)。
無論之後的森林古猿、臘瑪古猿,其實都是在強化“抓握”這個金手指而已。
而這個金手指還有一個作用,就是促進了大腦的發育。早在人類遠祖進化爲初始全颌魚的時候,因爲下颌的出現,咽腔縮小爲管道,給大腦的發育提供了空間。隨後長達數億年,到人類時,大腦已經占據了身體最大的比率。
800萬年前——大猩猩與我們分道揚镳。
700萬年前——乍得沙赫人,是我們和黑猩猩的近親。
乍得沙赫人,拇指還很短。此時的類人猿一支保留了原來的大拇指,演化成爲了今天的黑猩猩。

一支把大拇指進化得更長了,他們就是南方古猿。
大拇指更長,更加靈活,更加富有抓握能力,所以南方古猿下地幹活了(通常說法是,森林古猿走出叢林,進化爲南方古猿)。
爲了能夠驅動靈活雙手,芯片必須再度升級(腦容量的提升)。
想想看,占據如此多處理資源的雙手,不提升芯片怎麽得行?

芯片提升,然後和語言方面(族群的發展,社交發展,有了語言的需求)又互相促進,芯片再度提升。
600萬年前——黑猩猩與我們分道揚镳。
580萬年前——黑猩猩分支地猿。

黑猩猩的大腦與人類的大腦極其的相似,它們幾乎相當于縮小版的人類大腦:

現代人腦容量1500ml左右,早期人種腦容量700-1000ml不等,而南方古猿的腦容量只有450-530ml,猴子的腦容量低到200ml。

300萬年前——南方古猿,腦容量500ml,常被恐貓捕食。
人類在近300萬年內,腦容量飛速增加,這是怎麽做到的?
2020年的一份研究論文顯示,人類與黑猩猩的差異,並沒有人們想象中的那麽大。
英國劍橋大學醫學研究理事會分子生物學實驗室Madeline A. Lancaster教授的的研究團隊,發現了一個新的分子開關——ZEB2 基因[9]。
這個開關,可以實現人與黑猩猩大腦的“無縫切換”:
人類神經祖細胞成熟時間在7天左右,而黑猩猩是5天左右。
對黑猩猩的大腦中的ZEB2 基因進行延遲激活,就可以令黑猩猩的大腦進行“充分”的發育,最終長得和人腦相似。而人類的ZEB2 基因被提前激活,則可以導致大腦的發育“不夠充分”,從而最終和黑猩猩的大腦相似。

即便不考慮ZEB2 基因,黑猩猩的大腦也更像是縮小版的人類大腦,會經曆幾乎一模一樣的發育過程:
原始神經胚形成→前腦發育→神經細胞增殖分化→突觸連結、神經回路建立→神經髓鞘化
人類與黑猩猩屬的基因差異只有1.3%,可能相當多的性狀差異,實際都是相應分子開關所決定的。
人類的演化和地球環境、氣候的改變息息相關。
200~300萬年前。
地球進入了第四次冰期,惡劣的氣候,給人類演化增加了史無前例的沉重砝碼。
非洲環境變得更加的惡劣,不僅幹旱,還越來越寒冷,食物越來越少。大量的人類和動物在這一時期滅絕。
ZEB2基因被延遲激活, 不僅腦容量開始提升,而且童年時間也開始延長,以令大腦進行充分發育。
2006年,自然雜志發表了一篇關于南方古猿的文章,詳細介紹了年僅2歲零4個月的迪基卡小孩(Dikika child)[10]
這是迪基卡小孩的大腦與黑猩猩大腦的對比:
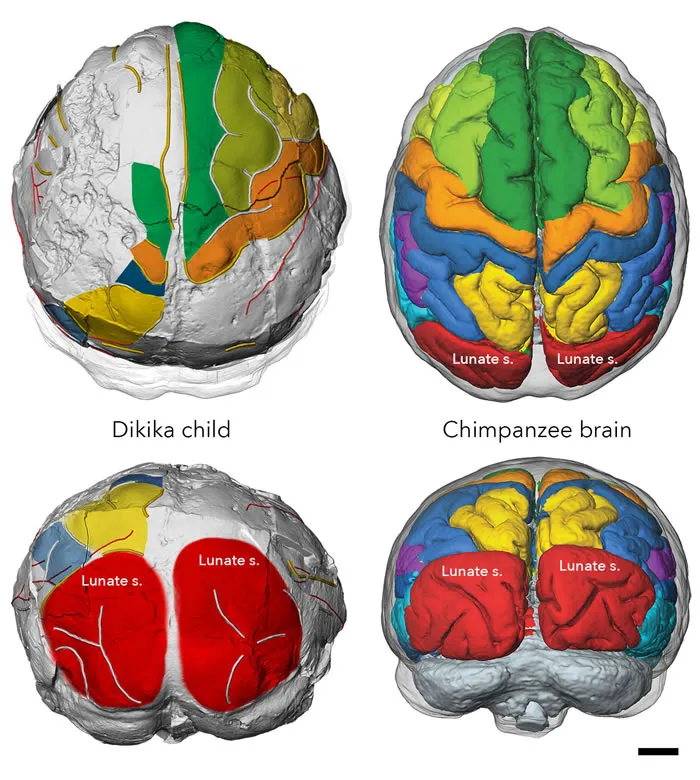
雖然迪基小孩大腦後側枕骨位置有專門負責視覺功能的溝槽,它的腦容量僅僅只有275毫升,是成年南方古猿成年個體的70%,相同年齡的黑猩猩,腦容量能達到成年個體的85%。
也就是說相比起黑猩猩,南方古猿幼年個體的腦發育速度明顯變慢了。
更加漫長的童年,在南方古猿身上便已經出現。[11]學習的更多知識和經驗, 幫助他們更加適應惡劣的環境。
隨著南方古猿活動區域的拓展,不同種群的産生是自然而然的事情。
但活動區域過大,必然産生物理隔離,由此形成生殖隔離,于是南方古猿演化出了不同的種屬。
例如阿法南方古猿、羚羊河南方古猿、 非洲南方古猿、驚奇南方古猿……
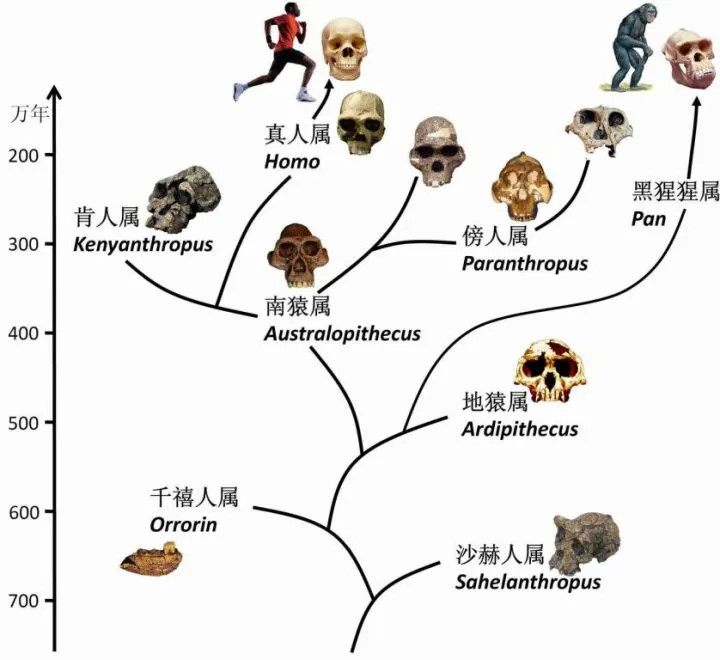
演化出直立人之後不久,南方古猿還是滅絕了。
要知道南方古猿的身高也就一米出頭,而直立人爲主的真人屬身高普遍達到1.5m,長高了近半個身子。
這裏可以看到一個十分殘酷的事實:個子矮小的古猿人,以及智力不夠高的,幾乎都在殘酷的自然之下系數滅絕。
也正是因爲殘酷的競爭,直立人大概就已經走遍了全球大部分地區(所謂第一次走出非洲)。
然而除了寒冷也就罷了,當時的非洲氣候,還出現周期性的波動。
在20萬年的時間內,曾出現過五六次森林到草原的變化過程。
惡劣的氣候,逼迫人類走遍踏入不同的大洲,但也因此出現地緣隔離,誕生了越來越多的人種。例如能人、樹居人、盧道夫人、匠人、直立人等等。

180萬年前——直立人,演化出了標槍投擲能力,腦容量1000ml,走向了食物鏈頂峰。
直立人從180萬年前走出非洲,而現代人是20萬年前走出非洲,直立人和現代人是完全不同的兩支人類。兩支人類都演化出了豐富的亞種。
150萬年前——匠人。
120萬年前——先驅人。

60萬年前——羅德西亞人、海德堡人,或爲智人和尼安德特人的遠祖近親。

此時人類腦容量已達甚至超過現代人水平的1400ml。
25萬年前——智人祖先出現。
智人的演化,實際從120萬年就已經開始,在40萬年前,才和尼安德特人的祖先分道揚镳。
16萬年前——長者智人(早期智人)。

7萬年前——走出非洲的現代人和尼安德特人發生過短暫的基因交流,所以除了非洲之外的現代人基因中,存在1%-4%的尼安德特人基因。此時,智人族群也一度衰落到2000人口規模(甚至有預估更少)。
5萬年前——現代智人(晚期智人)。
演化出來的現代人,生存能力無異于是最強大的。
一些人種自己滅絕了,一些人種則被現代人消滅了。
人類擁有極強的戰爭屬性,這或許是100多萬年前,殘酷的競爭已經銘刻到了基因之中。
我們作爲勝利者,除了一部分是自然滅絕的外,極有可能還有很多人種,是被我們親手埋葬(甚至吃掉)。
不僅僅尼安德特人,包括現代人種下的奧莫人、科羅馬農人、山頂洞人、柳江人也悉數滅絕。
人類演化過程中的殘酷,是無數現代人不可想象的。
以上便是用部分文獻支撐,用5000多字,盡可能詳細呈現的人類進化脈絡。
當然,如果反進化論的題主,你想反駁,歡迎拿出有力的論文證據對文中的關鍵信息,一個一個的對證。
如果你不想對證,且依舊質疑人類起源……
那——愛信不信。
參考^Lacalli T. The Middle Cambrian fossil Pikaia and the evolution of chordate swimming. Evodevo. 2012 Jul 6;3(1):12.^Xian-guang H, Aldridge RJ, Siveter DJ, Siveter DJ, Xiang-hong F. New evidence on the anatomy and phylogeny of the earliest vertebrates. Proc Biol Sci. 2002 Sep 22;269(1503):1865-9.^Lamsdell JC, Braddy SJ. Cope's Rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates. Biol Lett. 2010 Apr 23;6(2):265-9.^Zhu, M., Yu, X., Ahlberg, P. et al. A Silurian placoderm with osteichthyan-like marginal jaw bones. Nature 502, 188–193 (2013).^Simakov, O., Marlétaz, F., Yue, JX. et al. Deeply conserved synteny resolves early events in vertebrate evolution. Nat Ecol Evol 4, 820–830 (2020).^Irisarri, I., Baurain, D., Brinkmann, H. et al. Phylotranscriptomic consolidation of the jawed vertebrate timetree. Nat Ecol Evol 1, 1370–1378 (2017).^Zhu, Ya., Li, Q., Lu, J. et al. The oldest complete jawed vertebrates from the early Silurian of China. Nature 609, 954–958 (2022).^Gai, Z., Li, Q., Ferrón, H.G. et al. Galeaspid anatomy and the origin of vertebrate paired appendages. Nature 609, 959–963 (2022).^Benito-Kwiecinski S , Giandomenico S L , Sutcliffe M , et al. An early cell shape transition drives evolutionary expansion of the human forebrain. 2020.^[1] Alemseged Z , Spoor F , Kimbel W H , et al. A juvenile early hominin skeleton from Dikika, Ethiopia[J]. Nature, 2006, 443(7109):296-301.^skull scans reveal evolutionary secrets of fossil